|
ADVANCING MARINE
BIOTECHNOLOGY: USE OF OCS OIL PLATFORMS AS SUSTAINABLE SOURCES OF
MARINE NATURAL PRODUCTS
Principal Investigators:
Russell J. Schmitt,
Jenifer Dugan, Scott Hodges,
Robert Jacobs, Mark Page,
Leslie Wilson, and Steven Gaines
Summary of Research
The
potential for sustainable harvest of marine organisms that contain
valuable natural products, is an important question for the future
management and use of ocean. Many species of marine organisms
potentially contain compounds that could become valuable natural
products for a variety of applications including medicine, industry,
and food. However, harvest of these organisms in sufficient
quantities for applied uses could potentially have significant
ecological impacts, particularly for those species inhabiting natural
reefs.
 The
intertidal and subtidal portions of OCS offshore oil and gas production
platforms provide habitats for marine organisms, many of which also
occur on benthic and intertidal reefs. In waters off the coast of
California, platforms are known to provide habitat for a rich and
productive marine community containing over 50 species of algae and
invertebrates and reaching high biomass. The feasibility of
obtaining natural products having promising pharmaceutical applications
or other uses from species that grow attached to submerged portions of
offshore oil and gas production platforms is virtually untested.
Growth rates of some invertebrate species on platforms can be quite
high, for example, mussels and goose barnacles on oil platforms in the
region grow at rates equal to or higher that the highest reported
anywhere else in the world. Furthermore, many of the organisms
inhabiting oil platforms are encrusting forms which rely upon
antifouling compounds, rapid growth rates and alleopathic effects to
compete for and maintain space on hard substrates and avoid predation
(e.g., tunicates and sponges). Such characteristics have been
shown to be associated with compounds with potential pharmaceutical and
other applications. The
intertidal and subtidal portions of OCS offshore oil and gas production
platforms provide habitats for marine organisms, many of which also
occur on benthic and intertidal reefs. In waters off the coast of
California, platforms are known to provide habitat for a rich and
productive marine community containing over 50 species of algae and
invertebrates and reaching high biomass. The feasibility of
obtaining natural products having promising pharmaceutical applications
or other uses from species that grow attached to submerged portions of
offshore oil and gas production platforms is virtually untested.
Growth rates of some invertebrate species on platforms can be quite
high, for example, mussels and goose barnacles on oil platforms in the
region grow at rates equal to or higher that the highest reported
anywhere else in the world. Furthermore, many of the organisms
inhabiting oil platforms are encrusting forms which rely upon
antifouling compounds, rapid growth rates and alleopathic effects to
compete for and maintain space on hard substrates and avoid predation
(e.g., tunicates and sponges). Such characteristics have been
shown to be associated with compounds with potential pharmaceutical and
other applications.
The
organisms growing on platforms are sacrificed periodically when
the platforms are cleaned for structural safety. Using this
untapped source of marine organisms for natural products research (and
perhaps eventual harvesting) could eliminate virtually all of the
direct ecological effects that accrue from harvesting on natural
reefs. For example, in the Southern California Bight, mussels for
human consumption are harvested on a sustained basis from offshore oil
platforms, raising the possibility that platform species with desired
natural products also could be 'cultured' with minimum impacts on
natural reefs. Further, organisms that naturally dwell at very
great depths occur within scuba diving depths on the offshore platforms
in our region. This unique situation provides us with remarkably
easy access to a suite of species that otherwise would be difficult and
expensive to examine, and raises the possibility that eventual
mariculture of these species on OCS oil platforms also may be
feasible.
Overall,
our objective is to investigate the feasibility of using OCS oil
platforms for the development of marine biotechnology.
Specifically, we plan to explore the use of organisms growing on OCS
oil platforms for the development of pharmacological compounds.
To realize these objectives we will integrate research across three
disciplinary areas here at UCSB: 1, community and population ecology of
reef organisms (Dr. Jenifer Dugan, Dr. Mark Page), 2, population
genetics and natal sources (Dr. Scott Hodges, Dr. Steven Gaines), and
3, pharmacology with an emphasis on natural products identification and
mechanisms of action (Dr. Robert Jacobs, Dr. Les Wilson). We
believe that the long-term success of using OCS oil platforms for
biotechnology development will require basic information derived from
each of these disciplinary areas. For this proposal, the major
objectives addressed in each of these areas are:
COMMUNITY
AND POPULATION ECOLOGY. To survey OCS oil platforms for
organisms with potential applications to biotechnology, particularly
those with pharmacological applications. To determine the factors
affecting the growth and distribution of these organisms on OCS oil
platforms.
POPULATION
GENETICS AND NATAL SOURCES. To generate molecular-genetic
markers for the accurate identification of species with potential
applications to biotechnology. To determine the degree of genetic
diversity of organisms growing on OCS oil platforms relative to
natural reefs. Identify sources of populations for organisms
growing on OCS oil platforms.
PHARMACOLOGY.
To identify novel compounds, particularly
eicosanoids and coumarins, affecting inflammation, wound healing or
cell division from organisms growing on OCS oil platforms. To
investigate the mechanisms of action of these compounds for the
potential development of new drug targets.
ECOLOGY:
Background
Interest in marine natural products continues to grow
worldwide. However, concern over the impact of the harvest of organisms
that contain these products on the environment has arisen because large
quantities of organisms are typically needed to extract a small amount
of a natural product. Harvest of organisms from man-made structures,
particularly oil and gas platforms, may alleviate impacts to natural
reefs as many species of invertebrates grow on these artificial
structures. Unfortunately, little information is available on the
distribution and abundance and dynamics of invertebrates on oil
platforms. To examine the possibility of using OCS oil platforms
as sustainable sources of, or as culturing sites for, invertebrates
with important marine natural products, we have: 1) investigated
spatial and temporal patterns in the distribution and abundance of
invertebrates on selected offshore oil platforms in the Santa Barbara
Channel, 2) explored whether the population dynamics (recruitment and
growth) of common invertebrates vary among platforms (both spatially
and temporally), and 3) examined the relationship between patterns of
distribution and abundance and recruitment found at the platforms, and
selected environmental factors (e.g., location, water temperature). Data collection on the Ecology component of
this project is complete and manuscripts are in preparation.
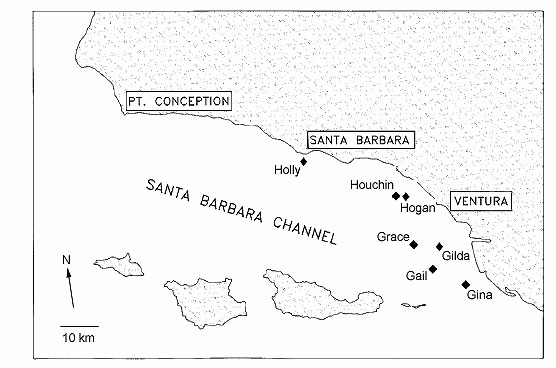
Study Sites
We conducted our research at seven
oil and gas platforms in the Santa Barbara Channel (Table 1, Fig.
1). The platforms are arranged along the channel from the
southeast to northwest in a region characterized by strong
environmental and biogeographic gradients.
Figure 1. Locations of oil platforms in
the Santa Barbara Channel involved in this study.
Table
1. Characteristics of
study platforms. Key to
abbreviations: Gi-Gina, Ga-Gail, Gil-Gilda, Gr-Grace, Hog-Hogan,
Hou-Houchin, Hol-Holly.
|
Variable
|
Gi
|
Ga
|
Gil
|
Gr
|
Hog
|
Hou
|
Hol
|
|
Distance
from shore (km)
|
5.0
|
13.2
|
11.9
|
14.4
|
5.1
|
7.0
|
2.9
|
|
Water
depth (m)
|
29
|
225
|
64
|
97
|
46
|
49
|
64
|
|
Age
(years from 2003)
|
23
|
16
|
22
|
23
|
35
|
34
|
37
|
|
Location
along channel (km)
|
0
|
12
|
15
|
19
|
33
|
36
|
65
|
|
Platform
size (m2 on bottom)
|
560
|
5600
|
2340
|
3120
|
1444
|
1444
|
1728
|
Preliminary
Results
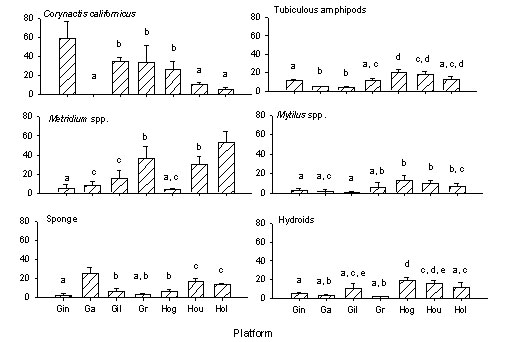
Spatial variation in platform invertebrate communities
Across all platforms, the most widely
distributed and abundant taxa, accounting for 83% of the total cover in
our photoquadrats, were anemones (e.g., Corynactis californicus,
Metridium sp.), tubiculous amphipods, (primarily Ericthonius
sp.), hydroids (Plumaria, Agalophenia), and sponges
(e.g., Haliclona spp., Halichondria panicea).
Other widespread taxa included mussels, (Mytilus californianus, M.
edulis), barnacles (Megabalanus californicus, Balanus
spp.), and tunicates (e.g., Styela montereyensis). Exotic
species were conspicuous on two platforms; the encrusting bryozoan, Watersipora
cucullata, was observed only on Platform Gilda and the anemone, Diadumene
sp. was recorded only on Platform Gail . Filamentous red algae
were the most widely distributed algal taxon. However, the cover
of algae was low (~5%) overall.
The structure of invertebrate communities varied greatly
among platforms (P<0.001, F=13.729, df=120, 1082.43, MANOVA:
Fig. 2). Anemones occurred in higher cover overall (up to 50 to
60%) than most other invertebrates, but the dominant species varied
with location. Corynactis californicnus was the dominant
anemone on platforms at southeast end of the channel (e.g., Gina, 59 ±
18%); cover of this anemone tended to be lower on platforms to the
northwest (e.g., 5 ± 2% at Holly). An exception to this pattern
occurred at Gail where mean cover of C. californicus was only 2
± 1% and the most abundant anemone was the exotic species, Diadumene
sp. (25%). In contrast, mean cover of Metridium sp.
was generally highest at the most northwest platforms (Holly, 51
± 13%) and lower on platforms to the southeast (Gina, 3 ± 2%). An
exception to this pattern was evident at Hogan where cover of Metridium
was only 2 ± 1%.
Tubiculous amphipods, hydroids, and mussels also
generally occurred in higher cover on platforms with increasing
distance along the channel from the southeast to the northwest (Fig.
2). For example, tubiculous amphipods occurred at 15 to 20% cover
on Hogan and Houchin, but <5% on Gail and Gilda. Highest cover
of mussels was recorded for Grace and Hogan (up to 25%) and lowest
cover at Gilda (<3%). In contrast, the cover of sponges was
more variable, with highest cover at Gail (up to 35%) and the two most
northerly platforms (Houchin, Holly). The bryozoan,
Watersipora cullcullata, occurred only on Gilda with mean cover of
41% (data not shown).
Figure 2. Comparison of the distribution and
abundance of the anemones, Corynactis californicus and Metridium
senile, sponges, tubiculous amphipods, mussels (Mytilus spp.), and
hydroids among study platforms.
Community patterns
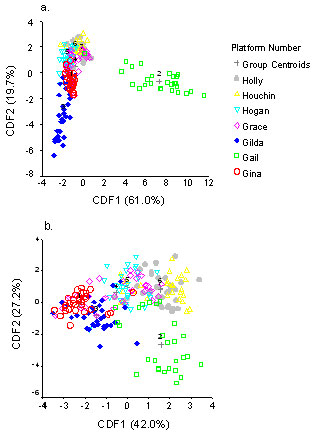
Discriminant Function Analysis (DFA) revealed that the
communities of Gail and Gilda were clearly different from the other
platforms, a pattern that can be attributed, in part, to the presence
of introduced species on these platforms (Fig. 3a).
Canonical Discriminant Functions (CDF) 1 and 2 explained 80% of the
variation in the data. Cover of the anemone, Diadumene
sp. was positively correlated (0.482) with CDF1, and an important
source of the separation of Gail from the other platforms along the
CDF1 axis. The negative correlation of cover of the bryozoan, Watersipora
cucullata, with CDF2 (-0.379) for Gilda contributed to the
separation of this platform from the others (Fig. 3a).
To explore the effect that the exotic species might have
on community patterns, we repeated the DFA, but excluded Diadumene
sp. and Watersipora cucullata from the analysis (Fig.
3b). The significant positive correlation of sponges (0.584) and
negative correlation of Corynactis californicus (-0.614) with
CDF1 contributed to the separation of all platforms except Gail along
the CFD1 axis. In contrast, the positive correlation of cover of Metridium
sp (0.605) and negative correlation of hydroids
(-0.428) with CDF2 contributed to the separation of Gail from the other
platforms along the CDF2 axis. Removal of W. cullculata
from the analysis reduced variability in the Gilda data and community
patterns at this platform tended to become more similar to those of the
next closest platform (Gina). In contrast, the structure of the
invertebrate community at Gail remained distinct from the other
platforms (Fig. 3b).
Figure 3. Results of Canonical Discriminant
Function Analysis of invertebrate communities on the seven study
platforms: a) all species, b) exotics species excluded.
Community patterns and
environmental variables
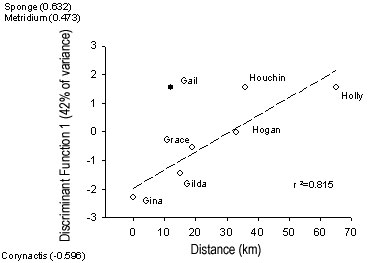
To explore relationships between community patterns and
environmental variables, we used the calculated values of CDF1 for each
platform (calculated including and excluding Diadumene sp. and Watersipora
cullculata), and the independent variables of location along the
channel, water depth, proximity to shore, and platform size (Table 1)
in stepwise multiple regression analysis. Prior to this analysis,
we tested for co-linearity among the independent variables. There
was a significant correlation between platform size and both water
depth (P<0.001, r = 0.974) and proximity to shore (P = 0.049,
r=0.758). However, depth and proximity to shore were not
significantly correlated (P>0.1). Therefore, we excluded
platform size from the analysis, but included water depth and distance
from shore. There was no relationship (P>0.1) between
variation in CDF1 and any of the independent variables if the data from
Gail were included in the analysis. If the data from Gail
were excluded from the analysis, variation in CDF1 was best explained
by location along the channel (P=0.014: Fig.4).
Figure 4. Relationship between canonical
discriminant function 1 and location of platforms along the Santa
Barbara Channel. R2 value calculated excluding data
from Platform Gail. Taxa most positively or negatively correlated
with CDF1 are also shown on the y-axis.
Measurements of water temperature during deployment of experimental
modules
The prevailing gradient in oceanographic conditions in
the Santa Barbara Channel is evident in satellite images showing the
intrusion of warm water into the channel from the south and cold water
from the west.
To examine variation in water temperature among
platforms, which could help to explain variation in community patterns,
a HOBO temperature logger was attached to one of the experimental
modules at each platform. Water temperature was recorded hourly,
with the loggers retrieved and downloaded at the end of each
experimental period. To compare temperatures among sites we
calculated the frequency of the number of hours at each water
temperature (Fig. 5).
Overall, the water temperatures at the three northern
platforms were similar to each other throughout the year.
Likewise, water temperatures of the four southern platforms were
similar during all seasons, except in the summer. During the
summer, one site (Gina) experienced large daily fluctuations in water
temperatures, resulting in a broad range of temperatures throughout
this season (Fig. 5). Such large temperature ranges were not
recorded at the other southern sites.
Figure 4. Water temperature frequencies by
degree hours for Summer 2001 at a) southern and b) northern platforms.
Figure 5. Distribution of water temperature,
expressed as percent of total number of hours at a given water
temperature, for the study platforms.
(4)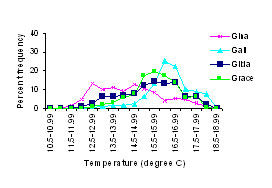 (5) (5)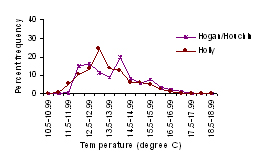
Recruitment Patterns and Oceanographic Factors
Recruitment data are useful for evaluating which
platforms may provide sustainable sources of marine natural
products. There were significant spatial and temporal differences
in patterns of recruitment of several taxa, although to varying degrees
(Fig. 6). For example, for some species, recruitment was higher
at the southern platforms (barnacles; Balanus trigonus
and B. regalis), while for others recruitment was
higher at the northern platforms (hydroids; Plumularia
sp.). Further, for some species recruitment was spatially limited
to just one (encrusting bryozoans; Watersipora cucullata)
or two platforms (tunicates; Diplosoma literianum).
Likewise, temporal patterns of recruitment varied among taxa, with
recruitment of some species occurring seasonally while for others it
was more continuous (e.g., tunicates).
Figure 6. Comparison of spatial and temporal
patterns of recruitment of four taxa, the barnacles, Balanus
trigonus, the hydroids, Plumularia sp., the encrusting
bryozoans, Watersipora cucullata and the tunicates, Diplosoma
listerianum, among study platforms.
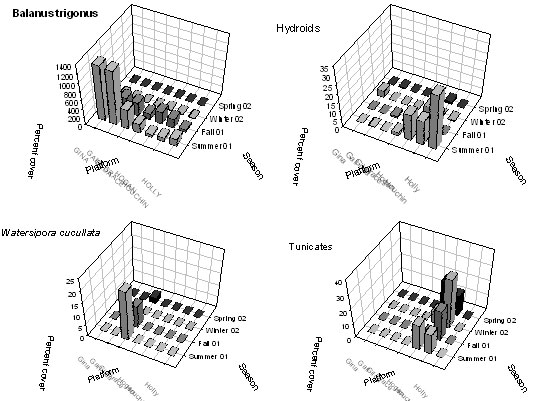
We used the nonparametric Spearman’s Rho (Rs) to explore
relationships between patterns of recruitment of selected invertebrates
onto plates at the study platforms and location in the Santa Barbara
Channel, platform depth, distance from shore and oceanographic factors
(Table 2). For the three barnacle species, there was a
significant effect of location, with higher recruitment densities
occurring at the southern compared to the northern platforms.
This pattern was consistent with predictions of oceanographic
conditions bringing warm water masses and the longer-lived planktonic
larvae of southern taxa into the channel. Recruitment density was
also associated with location along the channel for the hydroid, Plumularia
sp. For this species, recruitment was higher at the northern than
at the southern platforms.
In contrast, patterns of recruitment were variable for
the majority of invertebrate taxa with short-lived larvae or direct
development.
For most of these species, there was no relationship
between recruitment and location; many of these organisms recruited at
only a few platforms (e.g., Diplosoma listerianum) or a single
platform (Watersipora cucullata) where mature colonies occurred
in high abundance. This recruitment pattern is consistent with
the short larval development time and limited dispersal of these
species. Although there was an association between recruitment
and location for hydroids, their limited dispersal ability (crawl away
larvae) suggests that oceanographic factors associated with water
masses likely had little influence on transport of the hydroid larvae
and subsequent recruitment. Instead, biological interactions
(predation, competition) or other factors may have influenced the
recruitment patterns of this taxon.
Table 2. Relationships between recruitment of
selected invertebrates and location in the channel. Rs = Spearman
correlation coeffiencient. * < 0.05; ** < 0.01; *** < 0.001.
nsv = no significant variation in recruitment. nr=no recruitment
Species
Summer 2001 Fall
2001 Winter 2002
Spring 2002
Barnacles
Balanus trigonus
-0.857**
-0.893***
0.607 0.509
Megabalanus californicus
nsv
-0.321
nsv
-0.786*
Balanus regalis
nsv
nr
nsv
-0.821*
Encrusting
bryozoans
Watersipora
cucullata
-0.204
-0.204
nr
-0.204
Other encrusting bryozoans
nsv
-0.107
-0.054
0.071
Branching
bryozoans
Crisia complex/
Bugula
neritina
0.071
0.036
-0.286
0.029
Hydroids
Plumaria sp.
0.901***
nsv
0.056
0.089
Tunicates
Diplosoma listerianum
0.445
0.0490
0.0490
0.045
Our water temperature data provide support for the
hypothesis that oceanographic factors influenced recruitment patterns
in the summer for those species with longer-lived planktonic larvae, as
a gradient in water temperature occurred along the Santa Barbara
Channel during this season. In particular, warm water intrusion
was detected at the southern, but not at the northern platforms (Fig.
5). There was a significant correlation between barnacle (B.
trigonus) recruitment and water temperature in the
summer, both with (p = 0.05, Spearman’s Rho) and without (p = 0.0003)
the outlying data from Platform Gina (Fig. 7). This suggests that
larvae of warm water species could have been transported in these water
masses during the summer.
Figure 7. The relationship between recruitment of Balanus
trigonus and water temperature (mode) at each location. Open
circle = Platform Gina.
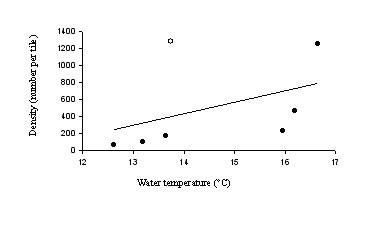
GENETICS:
We have focussed on determining the genetic variation
among populations of Bugula neritina. Primarily, we
analyzing DNA sequence variation for the mtDNA segment we have PCR
amplified. A preliminary analysis of samples that are clearly
closely related to the cryptic B. neritina species that harbors
the bacterium that produces Bryostatin-1 appear in Figure 1. This
analysis has two particularly interesting outcomes. First, it strongly
supports that we have identified a new cryptic species of B.
neritina. Second, it suggests that all of the samples from two OCS
oil platforms are members of a single clade and thus that colonization
of platforms may be a relatively rare event. We have also designed a
new pair of primers for amplification from the bacterial symbiont in
order to assess whether the new B. neritina clade also harbors
a unique lineage of symbionts (one that may produce a unique Bryostatin
compound).
Currently, we are
analyzing the mtDNA sequence data and will begin conducting the
bench-work to amplify and sequence DNA from the bacterial symbiont. Our
goal is to determine if there is a unique genetic lineage of the
bacterial symbiont in the new bryozoan lineage we have identified
(Figure 1).
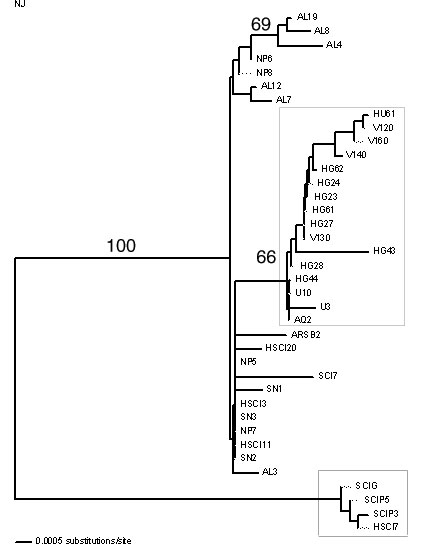
Figure 1.
Neighbor joining analysis of
DNA sequences from the 1.4 Kb mtDNA region of Bugula neritina.
Numbers above lines indicate bootstrap support (1000 replicates). The
large box encompasses the lineage containing samples from platform Hugo
(HG) and platform Houchin (HU). The smaller box encompasses the new
lineage of B. neritina found from samples from the Channel
Islands.
PHARMACOLOGY:
We are optimizing a number of organic extraction
procedures in order to obtain a favorable yield of pure extract from
Waterispora cucullata (WC). We confirmed that the activity was
retained by testing the pure compounds in the Sea Urchin Embryo assay.
<>Fresh samples were extracted from Waterispora
Cucullata (WC) and assayed on HeLa S3 cells for evaluation on cell
proliferation and viability.
Results
Inhibition of Cell Proliferation and Viability
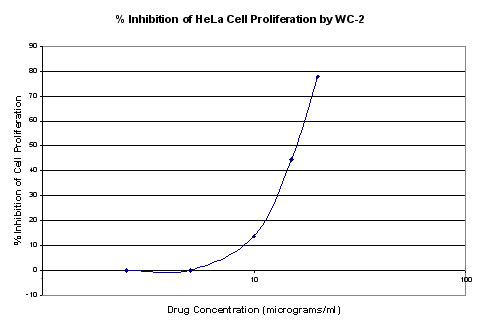
Plots were made of % inhibition of cell proliferation
(average of 2 samples) vs. drug concentration. Percent %
inhibition data are shown in figures 1 and 2. IC50 values were
approximated from plots and determined to be 12 μg/ml for WC1 and 16
μg/ml for WC2. Cell viabilities were determined for each drug
concentration by trypan blue dye exclusion and were found to be equal
or less than controls in all cases (control viability 98.9%)
Mitotic Index, Spindle and Chromosome Abnormalities
The mitotic index was determined from DAPI-stained
coverslips of untreated and drug treated samples. (Table 1
below). A 3 fold increase in mitotic index was seen in the two
highest WC1 samples but the mitotic index of the WC2 sample was
slightly below that of the control. A high percentage of mitotic
cells on the WC1 treated coverslips were noted to have abnormal
chromosome distributions (see attached PowerPoint Slide for
example). The most common abnormal cell type was a mitotic cell
with a distinguishable metaphase chromosome alignment but with 1 or
more chromosomes stuck at the pole. Several years ago Mary Ann
Jordan developed a classification scheme for these where she grouped
the abnormal cells as having either type I, type II or type III
spindles. (Jordan et al. 1992 J. Cell Science 102:401-16).
Type I spindles have elongated astral microtubules (we suspect most of
these were supposed to be kinetochore microtubules but never made the
right connections) and a metaphase chromosome distribution with one to
4 chromosomes at the pole. Type II spindles have a shorter than
normal interpolar distance (although still with long astral
microtubules) and more than 4 chromosomes at the poles but still have
recognizable metaphase accumulation of the majority of chromosomes at
the equator. Type III spindles form a ball of
chromosomes as a result of a collapse of the spindle into a monoastral
configuration in place of the normal bipolar orientation. We have
observed these types of cells as a result of treatments with drugs such
as the Vinca alkaloids, podophyllotoxin, and taxol type
compounds. This morphology is observed when nanomolar
concentrations of these drugs are used; concentrations that do not
alter microtubule polymer mass. From studies of interphase cells
treated with these concentrations of drugs we know that microtubule
dynamic instability is being suppressed by these drugs at these
concentrations.
Most of the abnormal cells in WC1 treated cells fell into
the category of type I and type II spindles. The proportion of
type I and type II mitotic spindles to total metaphase cells was
determined and is reported in Table 1. The abnormal mitotic cells
were rare in control populations and were not observed in WC2-treated
cells. Also notable was the lack of increased numbers of
multipolar spindles in WC1-treated cells (HeLa cells spontaneously
develop 1-2% of these as a proportion of total mitotic cells but no
increase in this number was seen in WC1 treated cells-data not
shown). Multipolar spindles are common in cells treated with
compounds like taxol and epothalone B, but not with Vincas and other
drugs that at higher concentrations cause microtubule
depolymerization. I think we can safely conclude that the
crude extracts contain at least one compound that promotes microtubule
stabilization in a manner similar to that of the Vinca alkaloids.
Table 1
|
Treatment
|
Mitotic
Index
|
Type
I spindles/total metaphase
|
Number
of cells counted
|
|
0.2%
DMSO
|
4.4
|
0.03
|
606
|
|
WC1
(40 μg/ml)
|
13.5
|
0.79
|
572
|
|
WC1
(20 μg/ml)
|
13.3
|
0.72
|
563
|
|
WC2
(20 μg/ml)
|
3.4
|
0
|
500
|
Figure 1
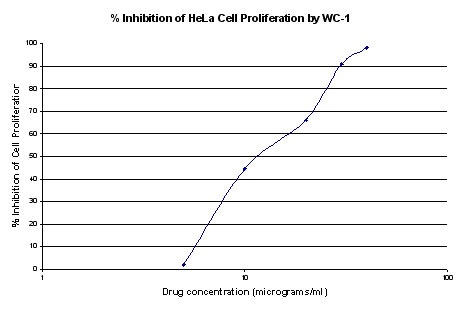
Figure 2
A graduate student , Daniel Day, is working in the
structure elucidation of the pure compound extracted from Waterispora
Cucullata (WC). After studying a number of established protocols we
were able to once again significantly increase the yield of pure
compound extracted. The methods and results are described in the
following paragraphs and figures.
The crude extract was prepared for thin layer
chromatography using 500 micrometer preparative silica gel plates with
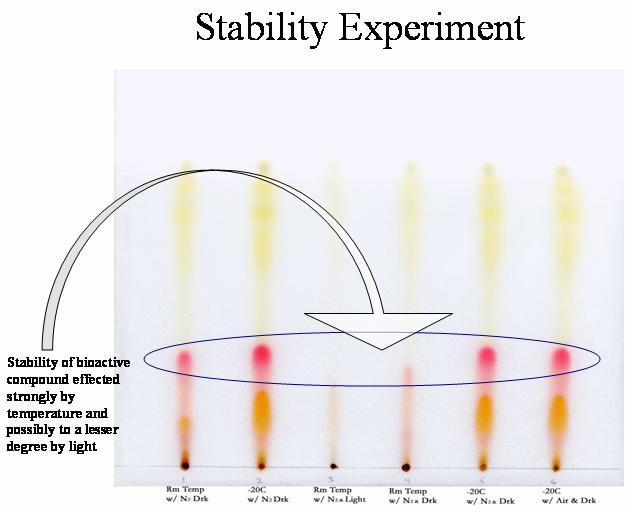
a mobile phase of 100% THF (Fig. 3). A recent experiment on
compound stability showed that the compound of interest (red compound)
is highly sensitive to temperature and slightly altered by light (Fig.
4).
Figure 3.

Figure 4.
3. Purification of the red band
After further purification with higher quantities
of starting material it was found that the previously described red
band was composed of 3 or more individual compounds including a unique
blue colored band which is highly unstable (Fig. 5).
Figure 5.
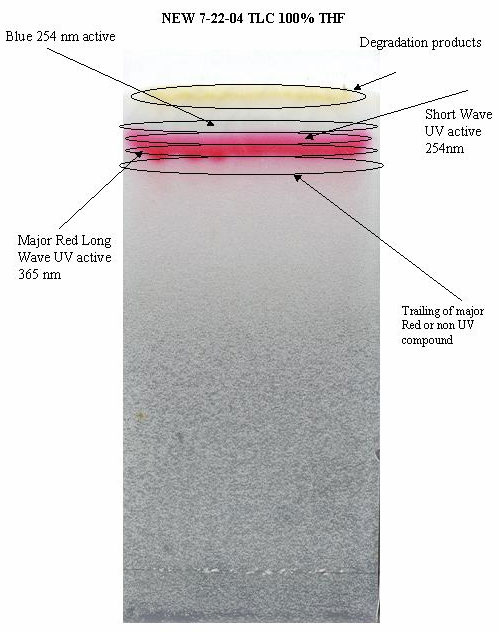
4. Activity assessment of purified bands
In order to verify that the newly purified
compounds (purified bands) retained biological activity a sea urchin
embryo assay was performed. All bands showed 100% activity in
inhibition of cell cleavage in the first cell division. At this
point we are focusing our attention on the major red band, which can be
seen on the previous TLC plate. Figure 6 shows a UV scan of the red
active compound and figure 7 shows a mass spectroscopy analysis with a
major peak at 311 m/z.
Figure 6.
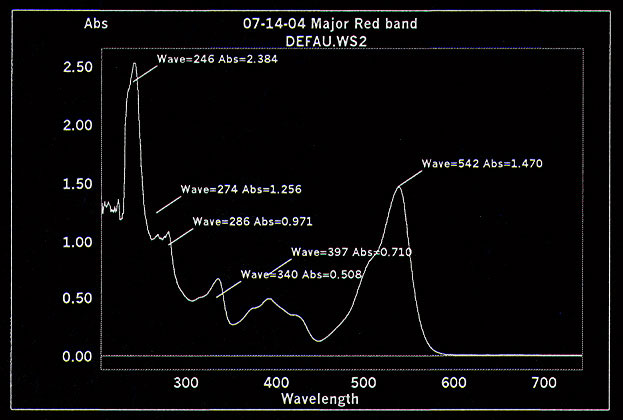
Figure 7.
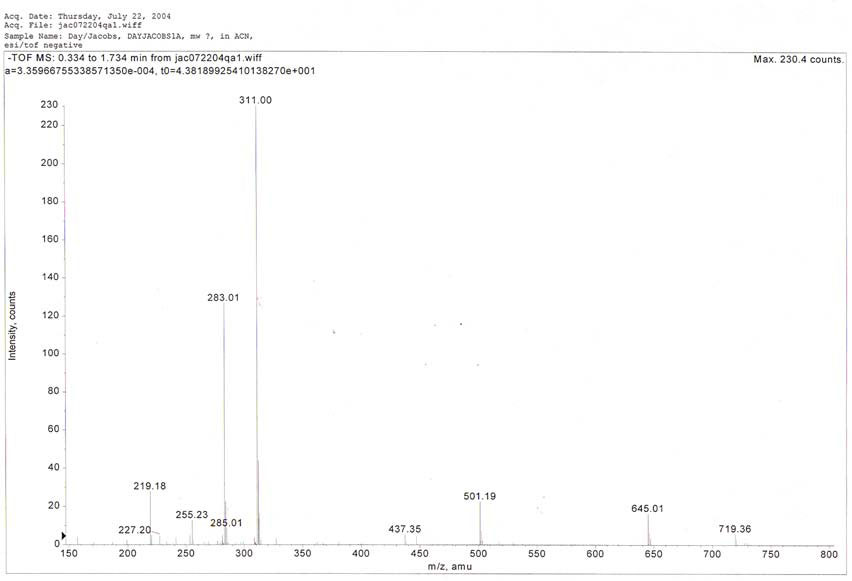
We are currently continuing analyses, including
more mass spectra, NMR, and elemental analysis in pursuit of a
structure for the compound, which we believe, is a novel compound.
|
[homepage]
|
[overview]
|
[researchers]
| [students
& staff]
|
[research] | [download reports]
|
|
